>>Terms like 'basal', 'early-diverging', and 'first-branching'
reflect persistent misconceptions about evolution and phylogenies
Why take the time to
blog about the issues with the use of the word "basal" and similar
terms?
This a good question because, indeed, many tree-thinking
papers have directly discussed misconceptions related to interpreting
phylogenies (e.g., Omland
et al. and Meir
et al.). In fact, several have tackled
issues surrounding the term "basal" specifically (see Krell
and Cranston and Crisp
and Cook). Given these efforts, I hoped that the use of this term (and the
associated misconceptions) would begin to erode. Unfortunately I feel as though, if anything,
the problem is becoming more widespread.
I think this is largely for several good reasons -- building phylogenies
is continually becoming easier, even for large datasets, and many researchers
from a range of fields are seeking to incorporate an evolutionary perspective
into their research. However, it's
important to point out that the misinterpretation of phylogenies is just as
common in evolutionary biology as in any other field. So in short, the “basal” problem is not going
to disappear without active efforts to teach tree-thinking to all biologists at
all stages.
So what is the
problem with 'basal', exactly?
The problem is that the term is used incorrectly and/or in
misleading ways in talks, papers, and proposals, roughly 90% of the time (by my
estimate). Moreover, the use of basal
and similar terms perpetuates a large suite of misconceptions about how
evolution works. So in order to
communicate effectively and accurately about evolution, we must also
communicate effectively and accurately about trees. As I have struggled to understand the desire
to describe some taxa as basal (or early-diverging or early-branching), I've
assembled a mental list of the various ideas that speakers and writers seem to
be aiming to communicate with the use of these terms. I've listed these below along with comments
about relevant misconceptions. This list
overlaps with what has been described in the publications listed above.
1) A basal species is
one that has given rise to another species, i.e. some ancestral lineage.
I think this use of the term is related to the misconception
that some living taxa are the ancestors of other living taxa. Unless something has changed with respect to
the space-time continuum, this is not possible.
The ancestors are lineages that are no longer present -- they are
represented in the tree as internal nodes and internal branches.
2) A basal taxon is one
that is older than other taxa in the tree.
If all of the taxa at the tips are extant (i.e., not
extinct), then all of them are the same age.
They all have the same root-to-tip distance in terms of time. In other
words, they have all evolved the same amount of time from the base of the tree.
(Note that this is a correct use of base -- the base is the earliest part of
the tree, the root, and time proceeds forward from that point.) It is worth noting that in molecular
phylogenies, some tips may be longer or shorter (i.e., the tree may not be ultrametric). This is due to a combination of stochasticity
in the substitution process as well as differences in substitution rates across
lineages. However, we would not say that the taxa on longer branches are
"more evolved" than the other taxa on the tree.
3) A basal taxon is
primitive morphologically or in some other sense.
All species, extant or extinct, possess a mixture of characteristics
that are, in cladistic terminology, ancestral (plesiomorphic) or derived (apomorphic)
relative to other species. For example,
in reptiles having scales and four limbs is the ancestral state. Snakes have
retained the ancestral state of having scales but have the derived state of no
limbs. This highlights the fact that no
species can be accurately described in evolutionary terms as 'primitive',
'ancestral', 'lower', or 'basal' any more than they can be described as
'derived', 'advanced', or 'higher'. In this sense, the tree-thinking view of
species diversity is rather egalitarian.
All of the species on earth have evolved the same amount of time from
the last
common ancestor of all life some 3 to 4 billion years ago, and their
diverse forms reflect the accumulation of changes during their unique pathways
along the tree to the present. It's worth noting that this misconception (that
not all species are equally 'evolved', or equally 'advanced') has been linked
to the history of progressive ideas in evolution, and specifically, the notion
that humans
sit at the apex of a ladder of life. This is exactly why terms like basal
are more than just poor wording; they perpetuate the incorrect interpretation
of phylogenies as ladders of progress. We can't expect to improve
understanding of the tree-like nature of evolution while continuing to use
misleading terminology.
4) Basal lineages sit
at the base of the tree or at the bottom of the tree diagram.
The observation that certain lineages are near the bottom of
a tree does not reflect any aspect of evolutionary history; it is simply a
reflection of the choices made in drawing the tree. These choices are generally
guided by aesthetic and didactic motivations.
That is, the tree is drawn to best communicate the results of the
phylogenetic analysis in a visually appealing way. The root could be towards
the top or towards the bottom, and the authors can rotate trees at nodes and
bend branches. None of these drawing
choices alters the relationships depicted in the tree. Thus, the two trees below communicate the
same phylogenetic information (e.g., lizards are more closely related to humans
that to frogs), despite the fact that the nodes have been rotated. This exercise makes it apparent why you
cannot learn interpret a phylogeny from the order of the tips, only from the
order of the nodes. For more practice in
reading trees without being distracted by tip order or tree format, look here
and here.

You may be wondering, so if the order across the tips can be
rotated without changing the tree, how do authors choose among possible rotated
versions? Since we read from left to right, it is common to show the 'focal'
taxon toward the right and if humans are in the tree, we are almost always put
in that prime position! Just take a look at most biology textbook depictions of
primate phylogeny, like this one.
Some of these drawing choices may also not be apparent to
novice users of phylogenetics software. Most inference software and tree
drawing programs will automatically 'ladderize'
trees, which places the sparsely sampled outgroups on the left of an upright
tree as above or on the top or bottom of a horizontal tree. Thus, the order of the tips that a program
produces is drawing convention and never the outcome of an analysis (e.g., you
could never say that 'the maximum likelihood analysis placed lemurs at the base
of the primate phylogeny').
So what should I say,
if not basal?
I realize that this whole post may be a big bummer. Especially if what you wanted from a
phylogeny was to learn which species were ancestral or which diverged first
(reminder, neither are possible or realistic, see above and below). Once you
have reconciled yourself with the fact that the ancestors are not among us,
here are some 100% not-confusing, evolutionarily-consistent, and still
interesting things you can say about a tree like the Platanthera orchid phylogeny below from this review
paper.

- The ancestors of extant Platanthera had nocturnally pollinated flowers. [Not, e.g., 'Nocturnally pollinated Platanthera are ancestral.' Because this could connote that some living Platanthera are ancestors of others. See space-time-continuum above]
- Diurnal pollination has arisen multiple times in Platanthera. [Not, e.g., 'Diurnally pollinated Platanhera are evolutionarily derived.' Because character states can be derived but taxa cannot be]
- The sonoharae-fuscescens-ussuriensis-japonica clade is sister to the rest of the genus Platanthera. [Not, e.g., 'The sonoharae-fuscescens-ussuriensis-japonica clade is basal to the rest of Platanthera.' or 'The sonoharae-fuscescens-ussuriensis-japonica group is an early diverging clade of Platanthera.' More on early-diverging below]
- The evolution of white flowers evolved after the transition to diurnal pollination in the clades that includes. P. blephariglottis and P. nivea.
- The basal nodes of the tree are reconstructed as nocturnally pollinated. [It is fine to describe the earliest nodes in the tree as basal because they did in fact occur earlier than the nodes towards the tips. Although personally, I prefer 'deeper' nodes.]
Notice that in this list, I use basal to refer to nodes,
sisters to refer to taxa, and ancestral/derived to refer to characters. My intuition is that many who use basal to
refer to taxa are actually most interested in characters (what did the ancestor
look like, not who was it exactly). So
if this is the case, talk about the characters! This is a good strategy for
avoiding tree mis-speak.
How about calling
some taxa 'early-diverging' or 'first-branching' instead of 'basal' or
'lower'? Doesn't this do the trick?
In short, no. These terms are equally uninformative, and
since many listeners will equate them with basal, equally misleading. I'll walk through an example that may help to
reveal why early-diverging is not meaningful.
Let's first consider a very simple tree with just two tips.

We would look at this tree and say there are two tips that
have diverged from a common ancestor and have evolved for the same period of
time since that split. We can add
another taxon to this tree.

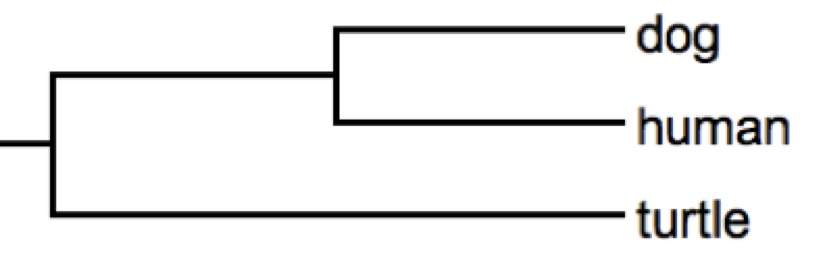
It is still true in this tree that the turtle and the human
have evolved the same amount of time since the earliest node in the tree. And so has the bird, remembering that its ancestry
includes the branch shared with the turtle.
Looking at this sort of topology, there is a tendency to call the branch
labeled human as 'early diverging', but of course the bird-turtle branch
diverged at the same moment. Thus, this
term seems to instead to be applied to whichever branch has given rise to fewer
descendants given the taxon sampling. Taxon sampling, like tree drawing, is a
choice, and I could instead have chosen a different set of three taxa, e.g.,
Calling the turtle an early-diverging amniote based on this
tree is just as odd as calling the human an early-diverging amniote based on
the previous tree. Even if we had all of
the amniotes sampled in this tree, one of the two branches arising from the
root node would be less speciose than the other. Tree drawing software will typically
ladderize the tree and thus would place the less speciose clade at the bottom
of the figure. This placement does not
making it earlier-diverging -- just as with any pair of sister taxa, the two
descendent branches emerged from the node at the same time and have evolved for
the same time to reach the present. This is not affected by how many times
those branches have split.
I have also seen a tendency for some to say, well, what I
mean by early-diverging or basal is indeed species-poor sister group, so as
long as that's clear to the audience, I will continue to use this term. My first response is that pointing out that
one group is species-poor is not an interesting observation by itself. All real trees are unbalanced and one of the
two branches descending from the root will almost always have more descendants.
Second, I think this choice of wording is simply too dangerous. Using terms like basal and early-diverging
carry strong connotations, and most audiences will assume taxa carrying this
label have retained more primitive characters or will fall into one of the many
misconceptions listed above.
Looking forward
So what do I hope to result from this long exposition on tree terminology? Most of all to have convinced you that it's not about the terms, it's about the ideas. Evolutionary biologists, me included, spend tremendous energy to learn about the history of life -- when groups diverged from each other, what changes occurred along the different branches, what factors may have caused these changes. But this effort is wasted if sloppy terminology allows the inferred history to be misconstrued as a ladder of progress, or yet another living fossil. I believe that we don't need such familiar and comfortable storytelling to make evolution interesting or relevant, to our peers or to the general public. The truth is that all living taxa have traversed fascinating paths to reach the present and all of their stories are worth telling.
So what do I hope to result from this long exposition on tree terminology? Most of all to have convinced you that it's not about the terms, it's about the ideas. Evolutionary biologists, me included, spend tremendous energy to learn about the history of life -- when groups diverged from each other, what changes occurred along the different branches, what factors may have caused these changes. But this effort is wasted if sloppy terminology allows the inferred history to be misconstrued as a ladder of progress, or yet another living fossil. I believe that we don't need such familiar and comfortable storytelling to make evolution interesting or relevant, to our peers or to the general public. The truth is that all living taxa have traversed fascinating paths to reach the present and all of their stories are worth telling.
**I'd like to acknowledge advice from Emily Sessa, Brian O'Meara, and Eric Schranz on this post, as well as helpful comments from Matt Hahn.
Thanks for this Blog post that help biologists keep in mind that there is no basal species !
ReplyDeleteAnd do not forget that there is no "living-fossil" tooo !
http://www.ncbi.nlm.nih.gov/pubmed/23382020
(sorry for advertising my own paper)
I agree that the term “basal” is sometimes used incorrectly, but that doesn’t mean that the term is wrong and cannot be used correctly. I find “basal” a very useful term when talking to colleagues about phylogenetic trees. Basal means “close to the base of the tree”. Most issues raised are related to correct ways of using “basal”:
ReplyDelete1) This is only a misconception when talking about extant species but that species give rise to other species is a central principle of evolutionary thinking. Those parent species can be called ancestral or basal. I don’t see a problem with that.
2) Extant taxa are not the same age. Some taxa are older, some are younger. Age is measured from the present to the time of origin or the age of the most recent common ancestor of the clade. Either way, most taxa differ in age. But I agree that it is incorrect to call an old taxon “basal”, juts because of its age.
4) The fish-branch is basal in relation to the other four terminal branches depicted because it branches off closer to the base of the tree. I don’t see any problem with that use of the world “basal.”
1) This is part of the 10%. In the vast majority of cases, basal is applied to extant taxa. Also, it is *very* hard (maybe impossible) to demonstrate that one species is the direct ancestor of another. Why not focus on the placement of the fossil taxon on the tree and its implications for evolutionary history instead?
ReplyDelete2) The ancestry of all extant taxa extends from the present to the root, giving all tips the same root-to-tip distance in terms of time. We humans sit on a tip, which tracing back, goes all the way to the common ancestor of all life, just like every other extant species. So all extant species have evolved for the same amount of time. We may chose to name some branches (and naming is often where problems arise), but the names are just labels.
I'd like to add that this is another case where focusing on characters can help. It would be inaccurate/misleading to say that fishes gave rise to tetrapods, but it's totally accurate to say that vertebrae (shared by fish and tetrapods) arose before limbs. Thus, while problems arise if we try to order extant taxa, we can talk about order of characters evolving without confusion.
4) The fish branch is not closer to the base. It is just as close as its sister clade (frog-lizard-mouse-human). You may be distracted by the number of nodes. See these great papers about 'node-counting' and why it's not useful in reading trees:
http://www.evolution.berkeley.edu/UToL/meisel10.pdf
https://www.nabt.org/websites/institution/File/pdfs/publications/abt/2007/069-07-0040.pdf
I appreciate the author's goals, among them being to encourage true "tree thinking" and to move readers away from the pre-Darwinian Scala Natura. However, the above prose introduces problems of its own in an attempt to conform to cladistic terminology. One example is the claim that all taxa are of equal age. (Is the taxon "Bacteria" the same age as the taxon "Mammalia"? Are species the same age as the higher level taxa in which we place them?) Likewise problematic is the insistence that living taxa cannot have given rise to other extant taxa. Clearly this does not apply to higher taxa; and even as applied to "species" the assertion is problematic, as many a commentator has noted.
ReplyDeleteWe would not say that a higher ranked taxon has given rise to a lower ranked taxon -- e.g. We would not say that Eukarya has given rise to Mammalia. Instead, Mammalia is a clade within Eukarya. It is nested within the larger clade -- this is the essence of tree structure. Still, naming is about our choices as taxonomists, not about biology. We decided where to put labels on the tree -- which clades we want to name and which not. Then assigning ranks to those names...that's a whole different thread.
DeleteThis comment has been removed by the author.
ReplyDelete1) Very hard to identify ancestral species in practice but very true that they existed.
ReplyDelete2) Agree if you measure "age" from the origin of life to the present. But the age of clades and lineages is also measured from their origin at a speciation event to the present, a much more useful measure in many instances.
4) My point is that the fish branch is closer to the base compared to any of the other terminal branches. Of course there are two basal sister clades in most cases. The point is that the origin of the fish branch lies at the base of the tree, and for that reason I would call it "basal". That tree is too cartoonish and incomplete to really talk about relationships among vertebrate groups, but fishes are basal in the sense just explained but rodents are not basal, because their origin is somewhere in the mammalian radiation, well above the base of the vertebrate tree.
If there were 100 kinds of fish in that tree (100 terminal fish branches rather than just the one shown), you wouldn't be calling fish basal. This is just our tendency to call species-poor branches basal. That one long branch misleads us into thinking that it is special. It is not special.
DeleteSantiago mentions the age of a taxon, and uses this as a justification for the use of the term basal. I want to come back and clarify why I think these are unrelated issues.
ReplyDeleteHow old is that taxon? If it is a clade, which I would hope, then the age can be attached to three alternative time points: the time when this clade diverged from its closes relative (its root age), the time when it acquired its most distinctive derived trait (its apomorphy age), and the time when it began to diversify into the distinct lineages that we have today (its crown age). Depending how long a stem lineage is (how closely related the clade is to other taxa that we know about), these three ages could be quite similar or quite different. Nonetheless, Santiago is quite correct that two clades can have very different ages: Bacteria is an older clade than Mammalia, by any of these ages.
I suspect that Santiago's justfication for wanting to call Bacteria more basal than Mammals is something like this: If we start from the root node and trace the lineage up towards these two clades, we cross the threshold “into” Bacteria earlier in time than we cross into Mammals. But, I would argue, and am confident that Stacey would agree, that this is irrelevant and a poor excuse for using the term “basal.”
To make the case, first consider the case where the two clades, the “basal” taxon and the “non-basal” taxon are sister to one another at the root node (“base”) of the tree. In that case the two clades share the same root age, so this cannot be the basis for claiming that one is older than the other. What if you consider one to have an older apomorphy or crown age than the other? You would be welcome to that conclusion, and could certainly communicate this to a fellow scientist, but it has nothing to do with the position of these clades on the tree. Therefore, using “basal” as a way to communicate that of two sister clades, one had a later radiation into its extant diversity (i.e., crown age) than the other is incorrect.
Now lets consider the case that the two clades you are naming are not actually sister to one another, but one is nested within the sister group of the other. “Bacteria" and “mammals" is an example of this. Comparing the age of these two clades might be interesting in some circumstances (e.g., as a step towards estimating the diversification rate). However, the label “basal” does a poor job communicating this since it focuses our attention, incorrectly, on tree topology rather than the (root or crown) age of those clades.
But, suppose I draw a tree which is pruned to only include bacteria and mammals, meaning that these clades would appear sister. Would it then be okay to call bacteria basal or early diverging? Again, the answer is no. Bear in mind that the clade that is sister to bacteria is not “mammals” but “archaea + eukarya.” It might be true that the “mammal” taxon is younger than “bacteria,” but this is really because mammals is (must be) younger than “archaea + eukarya,” the larger clade of which it is a part. So, in short, the clade age argument for using the term "basal" or "early-diverging" does not work.
You might read this as a rant from a cladist (not that I consider myself a "cladist”): a case of oppressive “phylogenetic correctness.” But before you do, it is a good idea to ask whether, actually, you think that a trout is more primitive than a human. If you do, then I would say you still have misconceptions about the structure of evolution writ large. If you do not, then I would urge you to drop the “basal” or “early-diverging” language to help your students and colleagues confront their own confusions about macroevolution.
Thank you, David, for these helpful and clear examples. I agree with your comments, and you are quite right that this discussion is not about which nodes we assign taxonomic names or how deep those nodes are -- it is about the misleading and inaccurate descriptors that get tacked on to those names (basal, early-diverging, etc.).
DeleteI disagree. This discussion is about clearly communicating concepts, but if one uses intentionally misleading trees, that is the antithesis of clear communication. If one creates a tree that is so pruned to only list bacteria and humans, or bacteria and mammals, one has created a tree in which one knowingly has created a misleading view in that bacteria and humans are not sister taxa. If we are to argue against the use of basal because it is not clear, then we also have to argue it based on trees which are not knowingly misleading and demonstrably incorrect. Accurate communication must begin with accurate information. In point of fact, in the tree of bacteria and humans, it is perfectly permissible to argue that bacteria is basal or early-diverging, because they in fact are. The entire bacterial clade has a much longer history than simply humans or mammals. They did branch off from the lineage leading to mammals far earlier than mammals became a distinct lineage. Thus, when looking at the whole tree, bacteria as a group are basal, even if its extant species may not be basal relative to humans or mammals. Basal is a relative term and can only be properly used when the groups are defined correctly.
DeleteAll of these arguments against the use of basal really hinge on using extant taxa that live at the same time. For those, I agree with you. However, when one is using extinct taxa, I think it is fair to use the term basal when appropriate. Extant organisms had to come from somewhere. Just because we cannot prove whether or not a taxon is ancestral to another doesn't mean we should not talk about it. No cladogram can be proven. They are hypotheses subject to further testing. Thus, referring to a taxon as basal is nothing more than a hypothesis subject to further testing in the same way as the entire cladogram.
Thank you for this and I will ashamedly cross out several lines of lecture notes. Now on your links...
ReplyDeleteI'm glad you found it helpful! There is plenty of mis-information in the literature and in textbooks, so you shouldn't feel too bad. As an instructor, you really have to go out of your way to find resources for teaching tree-thinking and heading off misconceptions. Good luck!
DeleteI couldn't resist communicating a nitpicking observation (although I am in full support of this post).
ReplyDeleteStacey states that rather than using "basal" nodes (when it is appropriate) she prefers "deeper" nodes.
In the graph literature the root of a graph lies at the top of the graph (i.e. "upside-down" from what biologists typically draw). Nodes closer to the root are thus "higher", not "deeper" (while nodes closer to the tips are "deeper", not "higher"). A tree is, of course, a graph, specifically a directed acyclic graph (DAG). Non-biologists are thus legitimately irritated that biologists use "higher" and "deeper" in diametrically opposite ways than everyone else!
Does it really matter that biologists use the terms this way? I think it does. Per Stacey's usage, "deeper" nodes confusingly correspond to "higher" taxa! Since ideally taxonomy will be phylogenetically-informed, I think it makes sense to make things consistent: "higher" taxa correspond to "higher" nodes. Then maybe the graph theorists will want to play with us!
Hi Joseph,
DeleteThat's a completely fair comment. As long as readers remember that time runs from the root towards the tip, then maybe earlier nodes is the best term. I'll keep this in mind next time I'm in front of a non-biologist, graphically minded audience!
p.s. I sincerely love many of the terms those folks prefer because they are so botanical -- leaves instead of tips and cherries for pairs of tips, e.g.,
http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.115.6021&rep=rep1&type=pdf
Just a comment about this, now that I have some time post-GSA, but you'll never get rid of this 'deeper' thing in paleontology because 'deeper' means 'further back in time' - as can be seen with the common use of 'deep time' in geology. And there's a real physical allusion here: geologic columns in their simplest form equate time with depth. Going 'deep' really means 'going back in time'.
DeleteSpeaking from a history of science perspective, it would be interesting to zoom forward several decades and see whether the institutions of math or of geology ultimately have a greater impact on the terms used by systematic biologists.
ReplyDeleteThe use of correct terms is fundamental for science reports. Nevertheless, the ancestor is not among us - is wrong! Let's talk about viral phylogeny - take a look - Viral phylogeny in court: the unusual case of the Valencian anesthetist - BMC Biology201311:83
DOI: 10.1186/1741-7007-11-83, and related article.
Besides, - it is important to think about the use of OUTGROPS to interpret a tree diagram. Using correct OUTGROUPS the tree will be displayed from the most divergent to the less divergent - and then if fossil records are available the hypothesis about ancestor and derived lineages gets stronger...
Hi Francisco,
DeleteI found the figures in that original article kind of hard to read, but I'll just say that viral trees work like any other trees. From the abstract, the case appears like other famous applications of phylogenetics for forensics, e.g. the gastroenterologist HIV case:
http://www.pnas.org/content/99/22/14292.full
The authors use patterns of relatedness among samples to test hypotheses about the source (in this case, showing that the victim sequences were nested among putative source sequences, providing very strong evidence against the gastroenterologist). But the same things I said about the trees apply here. The viral sequences kept evolving from the point of common ancestry between the victim and the source such that none of them were identical to each other or to the ancestral sequences. Of course, if you could have sampled back at the exact time of transmission (and if that transmitted sample was homogenous), you could have captured the ancestral sequence. But regardless, the patterns of variation among the victim and putative source sequences are sufficient to make the case, despite the accumulation of changes since the common ancestral sequence.
I'm not clear about your thinking on the outgroups, but I'll just add that those again are a choice (a part of taxon sampling). So I'd hesitate to call any particular sample of outgroups 'correct'.
I don’t think the term ‘basal’ is going to or should disappear because it has its reasonable uses. Brief quotes from two of the references you cited highlight the problem* and when it is useful to use ‘basal’:
ReplyDeleteCrisp and Cook 2006:
“Misinterpretation becomes evident when authors use the terms ‘basal’ or ‘early diverging’ to refer to extant taxa.”
Krell and Cranston 2004:
“Nodes or branchings near the base are basal nodes or basal branchings. The ‘basal branch’ is the branch between the most basal node (the last common ancestor of the members of the study group) and the root (Kitching et al., 1998: 200). A ‘basal clade’ is a part of the tree ending at a node before two or more terminal taxa. A ‘basal taxon’ is a (hypothetical) ancestral species, a species of the stem line, not a terminal taxon. All other use of the notation ‘basal’ is incorrect and misleading.”
*luckily I stopped myself from saying they highlight the 'root' of the problem
I use 'base of the tree' and 'basal nodes' in my blog as an attempt to show these reasonable and informative uses. But as I said, in my experience, the term basal usually misapplied (either to taxa / lineages or branches that are not actually an internal branch near the root). These are the uses that carry strong connotations (e.g. about some taxa being 'lower' or 'less evolved') and that allow ladder-of-progress misconceptions to persist. I certainly don't expect basal to disappear -- what I'm trying to do is promote ways of talking about phylogenies that accurately communicate the nature of evolution.
DeleteIndividual basal taxa/clades, certainly a no-no, but for the 10%, the "basal grade": paraphyletic assemblages of lineages that exhibit symplesiomorphies compared to derived nested clades. Or do we just have to avoid particular terms, useful or not, because they have been poisoned by inappropriate use?
ReplyDeleteHi Michael,
ReplyDeleteI think the issue with 'basal grade' is that, if we were to imagine drawing a box around that paraphyletic group, the box would include things that we could meaningfully call basal (basal nodes) as well as things that we could not (taxa). So in my view, it's safer to stick with grade or paraphyletic group without the 'basal' descriptor. Also, as you allude to, the reason we want to talk about these groups is often because they retain some character state of interest. Fortunately, it's easier to talk accurately about the characters (free petals are the ancestral state in angiosperms or complete metamorphosis is the derived state in insects). We can also discuss about which taxa possess ancestral or derived states for different characters, without calling the taxa themselves basal or derived. So, personally, I've never found 'grade' a very useful term anyway.
p.s. The term 'grade' also has a history of use in ladder-thinking in the context of 'evolutionary improvement' so maybe that's one more reason to avoid it.. See http://www.nature.com/nature/journal/v180/n4584/pdf/180454a0.pdf
Stacey, You might like this example of the 90%, from recent news story on blindsnakes:
ReplyDelete"This is an unusual feeding behaviour for a snake and was completely unexpected, says lead author of the study Takafumi Mizuno of the Kyoto Institute of Technology in Japan.
"Taken together with brief reports from 1950s and 1960s on similar behaviours in other blindsnakes, our finding implies that prey-dismembering behaviour may be widely used in poorly-known basal [ancient] snakes," he adds."
Ouch..
DeleteA colleague pointed me to this really helpful guide for talking about trees:
ReplyDeleteZachos. 2016. Tree thinking and species delimitation: Guidelines for taxonomy and phylogenetic terminology. Mamm. Biol. 81:185.
http://www.sciencedirect.com/science/article/pii/S1616504715001007
My favorite line: "There is branching, but no branching off"
I don't mean to dissect your comment too much, but I think this statement about 'no branching off' from Zachos is really problematic, and I feel necessary to explain why. (This is parallel but not directly connected to 'basal'.)
DeleteMainly, this statement assumes that there is no such thing as what paleontologists refer to as budding cladogenesis, at least an observable form of that process. Even if you ignore all the statistical evidence for 'budding' dynamics in paleobiology (i.e. that branching-off must be occurring during the duration of an observed morphospecies in the fossil record), it gets pretty close to being directly observable in the microfossil record. And, from my perspective, its a really troubling statement to make because micropaleontologists have been historically slow to adopt quantitative phylogenetics (i.e most of their phylogenies are line-drawings); we don't need to give them more ammunition to ignore phylogenetics by claiming the dynamics they see can't be real (and I think they are probably mostly real). I've even seen the inability to account for ancestor-descendant relationships cited in writing as a reason to avoid quantitative phylogenetics in that field.
I just really wish that the phylogenetics community would be embracing enough of the dyanmics of paleontological data that we wouldn't conversations like this, where we make sweeping statements that are so deeply from an ultrametric no-fossil perspective. I really think that when we talk about this issue, we should think about whether what we're saying applies equally to a phylogenies of reticulating prokaryotes, fossil-only trilobites, fossil-rich foraminifera, some-fossil canids, and orchids with no fossils (just theoretical examples... maybe there are orchid phylogenies with fossils). We should qualify our statements if they don't apply equally across that range, rather than making generalized statements that only apply to what we think are typical phylogenies.
Hi David,
DeleteI think the phenomenon of budding cladogenesis (when two species diverge, and one remains relatively static while the other evolves a new form) is interesting and probably widespread. Not having a paleo background, I am not super familiar with the fossil literature on it, but perhaps you can share your favorite paper? Anyway, the reason I would still resist saying branching (or budding) 'off', is some suggestion that the more morphologically static sister lineage is not evolving. Perhaps its form is relatively static, but of course it is continuing to accumulate mutations and is likely evolving in ways unseen in the fossil record (e.g. biochemically, physiologically). Also, branching 'off' may give the notion, after the split of the two species, the static one remains the ancestor of the morphologically novel form. This would defy time and instead the correct description is that the two share a common ancestor (before their split) and that the static one has retained the morphology of that ancestor.
With regard to your more general statement, I also think it would be great to have better integration of paleontological data with molecular phylogenetics and it seems like there is a great deal of movement in that direction (e.g. tip-dating). Maybe some have already done this, but it seems like these newer disparification methods are very amenable to integrating morphological data from extinct and extant lineages. Also have folks published traitgrams with mix of extinct and extant lineages? Seems like that would be an obvious way to visualize a Gouldian Christmas tree.
With respect to your comment about 'applying equally' to different groups of organisms, I think these phylogenetic terms and concepts do. Reticulation can involve any lineage (i.e., it's not a just a challenge for phylogenetics of prokaryotes), and I think as we collect more molecular data, we'll increasingly find out just how broad and deep it is. Still, we have long known that signal of a divergent, branching history is abundant in molecular and morphological data across clades (e.g., http://www.botany.wisc.edu/courses/botany_940/06EvidEvol/papers/Archie1989.pdf), showing that the movement of genes among lineages has not erased the underlying tree. (And btw, there is a nice Orchid fossil pollinarium that has been used for calibration, http://www.nature.com/nature/journal/v448/n7157/full/nature06039.html). Some of these patterns (whether budding cladogenesis or reticulation) may be easier to quantify and study in some lineages than others, but I'm not ready to say that the terms/concepts don't apply. I think that the more we maintain open lines of communication among people working on different groups, at different timescales and with different approaches, the better our chances of discovering general features of the evolutionary process.
Thank you for posting this blog. I have learned a great deal from it and the comments. However, I have one concept that keeps nagging at me. It has to do when the terms “more” or “less” are used in front of basal (previous comments addressed this, but I am still confused). Is there an appropriate use of the terms “more basal” or “less basal”?
ReplyDeleteAs you might guess, in my view, adding "more" or "less" is not informative since I don't think the term itself (in the way it is typically used) is informative. I think the "more" or "less" usually goes with issue (4) in the blogpost. So in this case, if it's being used to mean towards the bottom of the diagram, then the person might want to say how far towards the bottom the taxon sits. But since all nodes can be rotated, making 'bottomness' an artifact of how the tree is drawn, the 'more' or 'less' only piles on to the confusion.
DeleteAbsolutely love the content that you share with us. It's so easy to understand and provide knowledge as well, thank you so very much for sharing such content. Looking forward to read your other post.
ReplyDeleteDating Older Women
Romantic Wedding Couple Poses
What Unconditional Love Means
friendship is better than relationship
This post is extremely radiant. I extremely like this post. It is outstanding amongst other posts that I’ve read in quite a while. Much obliged for this better than average post. I truly value it! wedding venues near georgetown texas
ReplyDelete